فایده
مرگ
قرنهاست، از زمان ارسطو،
دانشمندان و فلاسفه در تلاش برای حل این معما بودهاند. برای نمونه، لوکرتیوس1
شاعر و فیلسوف رومی در کتاب «درباره طبیعت و اشیا»2 نوشته است که «پیری
و مرگ پدیدههایی
مفیدند؛ چون باعث میشوند که برای نسل بعدی جا باز شود (Bailey
1947)». چنین دیدگاهی
تا قرن بیستم نیز در میان زیستشناسان به قوت باقی مانده بود. برای نمونه، «آگوست وایسمن» زیستشناس نامدار آلمانی
که در قرن نوزدهم میزیست، مانند لوکرتیوس اعتقاد داشت که «انتخاب طبیعی ممکن است
موافق تکامل سازوکارهای مرگ عمل کرده باشد؛ چون مرگ با ایجاد فضا برای افراد جوان
و دارای توان تولیدمثلی بیشتر، بقای گونهها را تضمین میکند Weissmann
1891))».
توضیح تکاملی اقتصادی3 تر برای پیرشدن
باید براساس شایستگی و انتخاب فردی باشد، نه بر پایه انتخاب گروه. در دهههای ۱۹۴۰ و ۱۹۵۰ سه
زیستشناس
تکاملی، یعنی هالدین4، پیتر مداوار5 و جرج ویلیامز6
متوجه شدند که پیری به«نفع گونهها» تکامل نیافته است؛ بلکه پیری به این علت بهوجود میآید که عملکرد انتخاب
طبیعی (و شایستگی) در سن پیری کاهش مییابد و ناکارآمد میشود. بعدها ویلیام
هامیلتون7 و برایان چارلزورت8 در دهههای ۱۹۶۰ و ۱۹۷۰ اندیشه آنان را از نظر ریاضی تقویت
کردند و امروزه نیز درستی نظریه آنان بهطور تجربی معلوم شده است.
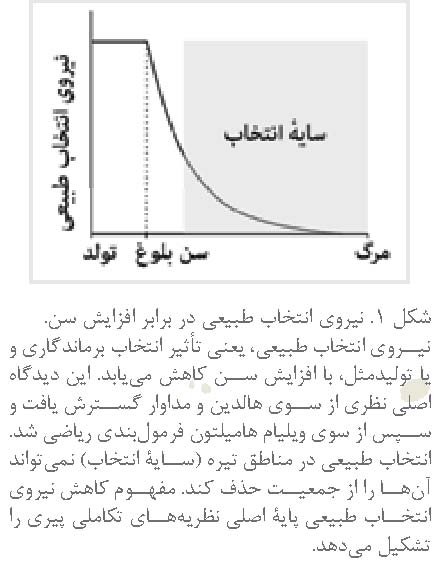
نیروی
انتخاب طبیعی با افزایش سن کاهش مییابد
دیدگاهی اصلی که باعث
شد مداوار و ویلیامز به نظریه تکاملی پیری برسند، بر این پایه استوار بود که با
پیشرفت سن، نیروی انتخاب طبیعی، یعنی میزان ماندگاری یا باروری، کاهش مییابد (نگاه کنید به Hamilton1966, Charlesworth 2000, Rose et al. 2007) (شکل۱). اولین بار فیشر9 در کتاب مشهورش، نظریه
انتخاب طبیعی10 (۱۹۳۰) تحلیل این موضوع را منتشر کرده بود؛ اما هالدین
(۱۹۴۱) و مداوار (۱۹۴۶، ۱۹۵۲) نیز بعداً به همین نتیجه رسیدند. هالدین (۱۹۴۱)
پیشنهاد کرد که کاهش نیروی انتخاب همراه با پیری، ممکن است شیوع نسبتاً بالای علل
غالب بیماری هانتینگتون را توضیح دهد: او استدلال کرد که چون بیماری هانتینگتون
معمولاً فقط در سنین بالای ۳۰ سالگی ظاهر میشود؛ بنابراین، انتخاب طبیعی نمیتوانسته است آن را در
اجدادی که در جمعیتهای پیشامدرن زندگی میکردند، حذف کند؛ زیرا بسیاری افراد پیش از آنکه بتوانند این
بیماری دوران مؤخر عمر را تجربه کرده باشند، مردهاند. بنابراین، انتخاب طبیعی اثری بر
این بیماری نداشته است.
مداوار (۱۹۴۶، ۱۹۵۲)
براساس اندیشههای
فیشر و هالدین اولین مدل کامل گرافیک تکامل پیری را توسعه داد. موضوع بحث مداوار
به این شرح است:
• اول، دنیای طبیعی برای بسیاری
جانداران، خطرناک است؛ چون پر است از رقبا، شکارچیان، عوامل بیماریزا، سوانح و تصادفات
و غیره. در نتیجه، در جمعیتهای طبیعی اغلب افراد قبل از اینکه پیر شوند و از سالخوردگی
رنج ببرند، میمیرند،
یا کشته میشوند:
بنابراین، احتمال اندکی وجود دارد که افراد در سنین پیری زنده بمانند و زادآوری
کنند (Moorad & Promislow 2010).
• دوم، تأثیر انتخاب طبیعی همراه با
افزایش سن کاهش مییابد (شکل ۱)، از اینرو انتخاب طبیعی عملکرد افراد را در
اواخر عمر نادیده میگیرد. در نتیجه، در سنین بالا انتخاب نمیتواند به نفع اثرهای سودمند، یا به زیان
اثرهای زیانآور
عمل کند. برای نمونه، اگر یک جهش مفید یا زیانآور بعد از توقف تولید مثل رخ دهد، بر
شایستگی11 (موفقیت تولید مثلی) تأثیر نخواهد داشت. با این حال، حتی اگر
یک جهش پیش از توقف تولید مثل هم رخ دهد، اثرهای آن ممکن است از سوی انتخاب قابل رؤیت نباشند؛ چون
اگر مرگومیر
بر اثر عوامل بیرونی، محیطی بالا باشد، ممکن است افراد پیش از آنکه آن جهش ژنی را
بیان کنند، بمیرند.
فرضیه
انباشتگی جهشها
مداوار (۱۹۴۶، ۱۹۵۲)
با توجه به منطق فوقالذکر، استدلال کرد که اگر اثرهای یک جهش زیانآور به سنین پیری
محدود شود، هنگامی که تولیدمثل متوقف میشود و امید چندانی به بقای بیشتر نمیرود، قبل از اینکه
اثرهای منفی در زندگی افراد حامل جهشهای منفی آشکار شوند، آن را به نسل بعدی منتقل کردهاند. در چنین وضعیتی،
انتخاب طبیعی از حذف چنین جهشهایی ناتوان و ضعیف است و در مسیر تکامل چنین جهشهای خنثایی در جمعیت
با رانش ژنی انباشته خواهند شد. این امر به نوبه خود منجر به تکامل پیری خواهد شد.
چنین فرایندی انباشتگی مداواری جهشها12 نامیده میشود (شکل ۲-الف). اثرهای چنین فرآیندی
تجمع جهشی بعد از تغییرات محیط تنها در سطح جاندار رخ میدهد؛ چون افرادی که دارای مرگومیر کمتر بیرونی
هستند (مانند کاهش شکار) و در نتیجه به سنی میرسند که در آن علائم پیری را بیان میکنند.
فرضیه
چندنمودی متضاد
جرج ویلیامز (۱۹۵۷)
با مقاله ارزشمند خود در زمینه تکامل اندیشه مداوار را یک گام به جلو برد. او
استدلال کرد اگر بپذیریم که انتخاب نمیتواند در رویارویی با اثرهای زیانآور پیری پیروز شود،
جهشها
یا اللهایی
ممکن است وجود داشته باشند که در سنین مختلف زندگی اثرهای متضاد یا چندگانه دارند:
تفاوتهای
ژنتیک که در سالهای
اولیه، هنگامی که انتخاب نیرومندتر است، بر شایستگی اثرهای مثبت دارند، در سنین
پیری که انتخاب ضعیف میشود، اثرهای زیانآور بر جای میگذارند. این اندیشه امروزه در مبحث
تکامل پیری «چندنمودی متضاد» نامیده میشود (Rose 1991,
Flatt & Promislow 2007, شکل ۲ ب). ویلیامز اشاره کرده است که اگر اثرهای
مثبت چنین جهشهایی
در جوانی بر اثرهای زیانآور آنها در پیری بچربد، چنین واریانتهایی مثبت خواهند بود و تقویت خواهند
شد. بنابراین، موجب تکامل پیری میشود. بنابراین، بر پایه فرضیه ویلیامز، تکامل پیری ممکن است در
جوانی محصول جانبی ضدسازشی انتخاب برای
ماندگاری و تولیدمثل باشد.

خلاصه
در این نوشته تفکر
زیستشناسان
تکاملی را در مورد تکامل پیری معرفی کردیم. امروزه، مشخص شده است که پیری فرایندی
نیست که انتخاب طبیعی آن را انتخاب کرده باشد، یا به نفع ماندگاری گونه باشد. بلکه
پیری به علت
عملکرد ضعیف انتخاب طبیعی و ناکارآمدی آن در زمان پیری است. برای کاهش نیروی انتخاب
طبیعی در سنین پیری دو فرضیه اصلی موجود است: انبـاشتگی جـهشها و چندنمودی متضاد. انباشتگی جهشها میگوید که پیری به این
علت است که انتخاب نمیتواند جهشهای زیانآور را که فقط در زمان پیری نمودار میشوند، حذف کند و طبق فرضیه چندنمودی
متضاد، پیری بهعنوان
محصول جانبی ضدسازشی برای افزایش شایستگی در جوانی روی میدهد. تأثیرات مثبت در جوانی از لحاظ
ژنتیک با اثرهای زیانآوری که باعث پیری میشوند، جفت میشوند. پیری آشکارا طول عمر را کوتاه میکند؛ اما طول عمر نیز
با انتخاب تعداد رو به افزایشی از رویدادهای تولید مثلی شکل میگیرد. بنابراین تکامل
دوره زندگی توازنی است بین آن دسته از عوامل انتخابی که دوره زادآوری را گسترش میدهند و اجزایی از مرگ
درونی که آن را کوتاه میکنند.
پینوشتها
1.
Lucretius
2.
De Rerum Natura
3.
parsimonious
4.
J.B.S. Haldane
5.
Peter B. Medawar
6.
George C. Williams
7.
William D. Hamilton
8.
Brian Charlesworth
9.
Fisher
10.
Theory of Natural Selection (1930)
11.
fitness
12.
Medawar's mutation accumulation
13.
antagonistic Pleiotropy
منبع
ترجمه
https://www.nature.com/scitable/knowledge/library/the-evolution-of-aging-23651151
منابع
و مراجع
Ackermann,
M. et al. Senescence in a bacterium with asymmetric division. Science 300,
1920–1920 (2003).
Ackermann,
M. et al. On the evolutionary origin of aging. Aging Cell 6, 235–244 (2007).
Austad,
S. N. & Fischer, K. E. Mammalian aging, metabolism, and ecology: Evidence
from the bats and marsupials. Journal of Gerontology 46, B47–B53 (1991).
Bailey,
C. Titi Lucreti Cari De Rerum Natura. Volume 3, Oxford, UK: Clarendon Press,
1947.
Blanco,
M. A. & Sherman, P. W. Maximum longevities of chemically protected and
non-protected fishes, reptiles, and amphibians support evolutionary hypotheses
of aging. Mechanisms of Ageing and Development 126, 794–803 (2005).
Bronikowski,
A. M. & Flatt, T. Aging and its demographic measurement. Nature Education
Knowledge 1, 3 (2011).
Charlesworth,
B. Evolution in Age-Structured Populations. Cambridge, UK: Cambridge University
Press, 1994.
Charlesworth,
B. Fisher, Medawar, Hamilton and the evolution of aging. Genetics 156, 927–931
(2000).
Charlesworth,
B. Patterns of age-specific means and genetic variances of mortality rates
predicted by the mutation accumulation theory of aging. Journal of Theoretical
Biology 210, 47–65 (2001).
Finch,
C. E. Longevity, Senescence and the Genome. Chicago, IL: University of Chicago
Press, 1990.
Fisher,
R. A. The Genetical Theory of Natural Selection. Oxford, UK: Clarendon Press,
1930.
Flatt,
T. Survival costs of reproduction in Drosophila. Experimental Gerontology, In
Press (2011).
Flatt,
T. & Promislow, D. E. L. Physiology: Still pondering an age-old question.
Science 318, 1255–1256 (2007).
Flatt,
T. & Schmidt, P. S. Integrating evolutionary and molecular genetics of
aging. Biochimica et Biophysica Acta 1790, 951–962 (2009).
Haldane,
J. B. S. New Paths in Genetics. London, UK: Allen & Unwin, 1941.
Hamilton,
W. D. The moulding of senescence by natural selection. Journal of Theoretical
Biology 12, 12–45 (1966).
Hughes,
K. A. & Reynolds, R. M. Evolutionary and mechanistic theories of aging.
Annual Review of Entomology 50, 421–445 (2005).
Hughes,
K. A. et al. A test of evolutionary theories of aging. Proceedings of the
National Academy of Sciences of the United States of America 99, 14286–14291
(2002).
Kirkwood,
T. B. L. Evolution of ageing. Nature 270, 301–304 (1977).
Luckinbill,
L. S. et al. Selection for delayed senescence in Drosophila melanogaster.
Evolution 38, 996–1003 (1984).
Medawar,
P. B. Old age and natural death. Modern Quarterly 1, 30–56 (1946).
Medawar,
P. B. An Unsolved Problem of Biology. London, UK: H. K. Lewis, 1952.
Moorad,
J. A. & Promislow, D. E. L. What can genetic variation tell us about the
evolution of senescence? Proceedings of the Royal Society B: Biological
Sciences 276, 2271–2278 (2009).
Moorad,
J. A. & Promislow, D. E. L. Evolution: Aging up a tree? Current Biology 20,
R406–R408 (2010).
Partridge,
L. & Barton, N. H. Optimality, mutation and the evolution of ageing. Nature
362, 305–311 (1993).
Promislow,
D. E. L. & Bronikowski, A. "Evolutionary genetics of senescence,"
in Evolutionary Genetics: Concepts and Case Studies, eds. C. W. Fox & J. B.
Wolf (Oxford University Press, 2006) 464–481.
Rauschert,
E. Survivorship curves. Nature Education Knowledge 1, 18 (2010).
Rose,
M. R. Laboratory evolution of postponed senescence in Drosophila melanogaster.
Evolution 38, 1004–1010 (1984).
Rose,
M. R. Evolutionary Biology of Aging. New York, NY: Oxford University Press,
1991.
Rose,
M. R. & Charlesworth, B. A test of evolutionary theories of senescence.
Nature 287, 141–142 (1980).
Rose,
M. R. et al. Hamilton's forces of natural selection after forty years.
Evolution 61, 1265–1276 (2007).
Shefferson,
R. P. Why are life histories so variable? Nature Education Knowledge 1, 1
(2010).
Stearns,
S. C. The Evolution of Life Histories. Oxford, UK: Oxford University Press,
1992.
Stearns,
S. C. et al. Experimental evolution of aging, growth, and reproduction in
fruitflies. Proceedings of the National Academy of Sciences of the United
States of America 97, 3309–3313 (2000).
Stewart,
E. J. et al. Aging and death in an organism that reproduces by morphologically
symmetric division. PLoS Biology 3, 295–300 (2005).
Weissmann,
A. Essays on Heredity. Oxford, UK: Clarendon Press, 1891.
Williams,
G. C. Pleiotropy, natural selection, and the evolution of senescence. Evolution
11, 398–411 (1957).